クジラとカバの意外な関係
*この記事は1999年、今は亡きサイアスなる雑誌に掲載されたものに補筆して、2002年頃にアップしたものです。まだ分子系統学の性質をちゃんと理解していないので(今も不十分かもしれませんが)、やや的外れなことも”もっともらしく”書いちゃってます。しかしまま掲載しておく次第。まあこういう時代もあったってことで。
要約:クジラは海で暮らす動物だが、同時に哺乳類でもある。そして哺乳類は陸上脊椎動物の1グループであり、陸上で進化してきた。つまり、クジラもそもそも陸上で生活する祖先から進化してきたのである。
そこで、これまでクジラの祖先がいかなる動物であったのか数多くの議論がおこなわれてきた。化石を研究する古生物学者たちは、絶滅した奇妙な有蹄類の一種、メソニックス類がクジラを生み出した動物群であろうと考えてきたし、それはほぼ間違いないだろうとも思われてきた。
一方、分子系統学は昔から違う結果を示してきたが、解析に問題点があることも指摘されてきた。しかし、近年になって開発された新しい手法、すなわち”レトロポゾンの挿入位置から系統を解析する”という方法はほとんどの問題点をクリアーする新しい手段である。東京大学の岡田典弘教授のチームはこの新しい手法によってクジラが偶蹄類(ウシやイノシシ、カバよりなる動物群)に含まれること、そして中でもカバにもっとも近縁であることを示した。この結論は分子系統学の最近の研究や、軟組織の特徴、そして幾つかの化石証拠によっても支持されている。
遺伝子上の奇妙な配列:レトロポゾンを用いた系統解析は何を明らかにするか??
海で暮らす哺乳類、クジラの祖先は陸上で暮らしていた。その祖先はどんな動物だったのだろう。それを知るためにこれまで多くの化石が採集され、また遺伝子の比較が行なわれてきた。
クジラと偶蹄類の類縁関係に新たな光りがあてられたのは1997年のことである。この年、東京工業大学、岡田教授の研究グループであった島村満たちは、レトロポゾンを用いることでクジラがカバとウシに非常に近い関係にあることを明らかにした。(Shimamura et al 97 )
そして、彼らが明らかにしたクジラとウシ、およびカバの意外な関係は、化石から考えられてきたこれまでの説とはまったく対立するものだった。
化石や解剖に基づく系統の推定と遺伝子を用いた系統の推定結果はしばしば食い違う。形態をもとに議論してきた何人かの科学者たちは疑問を持ったが、この研究結果が正しいことを暗示するような化石の発見も最近報告されている。そこで、クジラの祖先がどんな動物であったのか。その議論の歴史と岡田教授のチームの研究がそれにどんな影響を与えているのか、まずそれを見ていこう。
生物の進化の歴史を系統、それを調べる学問のことを系統学と呼ぶ。系統学には大きく2つの分析方法がある。ひとつは生物の形態を比較する分岐学(=Cladistics )で、もうひとつは遺伝子の比較に基づく分子系統学だ。これら2つの手法は相互に補完しあったり、あるいは対立したりしてきた。(長谷部 96 )そして、クジラの系統に関する議論では、対立の歴史であったと言えるだろう。
そもそもクジラが哺乳類であることが分かったのは17世紀末のことで、さらに19世紀末にはウシやウマ・ゾウといったひづめを持つ植物食の動物に近いことが示唆された。これらの動物は有蹄類と呼ばれ、植物を食べるほとんどの哺乳類はこの仲間であると考えられてきた。(有蹄類に関するもう少し詳しい内容は次ぎをクリックしてください>有蹄類へ)
19世紀末、クジラが有蹄類の仲間であると考えられた根拠は解剖学の視点に基づくもので、その後、近年になって確立した分子系統学もDNAを比較する立場からそのことを支持している。この点に関して両者の意見に違いは見られない。
しかし、系統のより細かい部分に関して見ると数十年の間、双方の科学者はお互いに対立する研究成果を出し続けてきた。
1950年、ボイドンとジェメロイは動物のタンパク質を比較して、クジラと偶蹄類は共通するタンパク質を、他の動物よりも多く持つことを明らかにした。これは間接的にDNAを比較したと考えてよい結果である。また、筋肉や内臓といった軟組織を比較した研究者たちもクジラと偶蹄類が多くの共通点を持つことを見つけている。こうしたことから著名なクジラ学者、アムステル大学のシュライパー教授はその著書の中で、クジラが偶蹄類に極めて近縁であることを指摘している。(シュライパー 65 )
だが、古生物学者たちの見解はいささか異なっていた。化石からのDNAの回収はほとんど不可能であり、軟組織、つまり肉や内臓が残ることもまずない。そこで古生物学者たちは骨や歯の形態に注目する。現代生きている現生種しか分析できない分子生物学者や解剖学者と違って絶滅種も比較できる彼らは異なる結論に達した。
1967年、ヴァンヴァーレンは絶滅した肉食性の有蹄類、メソニックス類の歯が原始的なクジラの歯と類似していることに気がつき、メソニックス類からクジラが進化してきたと考えた。(Van Valen 67)
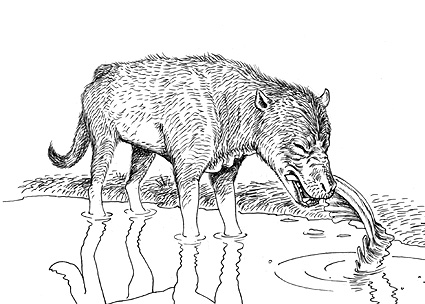
メソニックス類の一種、パキヤナエ。頑丈な顎と歯は堅い食物、例えば貝類や動物の骨を食べていたことを示す。イラストの中では動物の死体の脊椎骨からのびる肋骨をかじっている。
メソニックス類は歯が初期のクジラと非常に似ているので、古生物学者たちはメソニックス類からクジラが進化してきたと考えてきた。(この場合、メソニックス類はクジラの側系統群となり、人為分類群となる)
しかし、岡田研究チームの報告や近年の分子系統学の成果を踏まえるとメソニックス類がクジラに近縁であるかどうか疑問がもたれている。
例えばメソニックス類の歯は初期のクジラに似ているが、偶蹄類の歯とは似ていない。またかかとの骨、距骨の形態も偶蹄類と似ていない。またメソニックス類の距骨は初期のクジラのものとも似ていない。一方で軟組織や分子系統学、レトロポゾンの証拠はクジラと偶蹄類に類縁があることを明らかに示している。
困惑させられるのは、クジラと偶蹄類の類縁を支持する証拠は化石からもわずかながら見つかるが、クジラと偶蹄類とメソニックス、この3者を結び付ける証拠もわずかながらあるという事実かもしれない。(以上の事柄は詳しくは本文を参考のこと)
生物の特徴(形質)が矛盾した分布を示すのは普通に見られることだが、クジラと偶蹄類、メソニックス類に関しては錯綜の度合いがはなはだ激しいように見える。
さらに生物の形態から科学的に系統を推定する方法として、分岐学が受け入れられるようになると、この手法で解析が行なわれ、ヴァンヴァーレンの主張を裏付けるような分岐図(ここでは系統図とほぼ同じ意味で使う)が得られている。こうした分岐図が示すのは、クジラは有蹄類であるものの、偶蹄類に特に近い関係にはない、ということである。(Prothero et al 88)
一方で、分子生物学者たちは古生物学者が考えたものとは異なる系統図をつくり出し、さらに幾人かの研究者はクジラが偶蹄類に近いどころか、偶蹄類そのものであると結論した。
そのなかでも有名で、かつ最近のものが94年のグラウワーたちの報告で、クジラが偶蹄類に含まれること、また、偶蹄類の中でもウシに近いことをが報告されている(Graur & Higgins 94)。(ただし、この研究の解析に問題があることが指摘されている(長谷川&岸野 96 pp190~205)
また、同じ94年にアーウィンとアーナソンもクジラが偶蹄類に含まれること、さらにカバに極めて近いことを示唆している。
97年から始まる岡田教授の研究チームの成果は、以上で述べたクジラの起源にまつわる古生物学者と分子生物学者との長い対立に幕をひくものかもしれない。では、彼らが用いている”共通のレトロポゾンを発見する”という方法はいかなる方法なのだろうか。(岡田 97. 岡田 00 が参考になる)
レトロポゾン、この聞き慣れない単語は分子遺伝学の用語で、ある種の、DNAに逆流入した遺伝子配列のことを指す。そこでまずこの言葉から簡単に説明しよう。そのためには遺伝子の基本的なことを知らなければならない。
生物の身体のかなりの部分はタンパク質でできている。タンパク質は何種類ものアミノ酸から構成されていて、そのアミノ酸の種類と順番によって性質が決定づけられている。そして私達の細胞の中にあるDNAがアミノ酸の種類、その配列の順番を記録しているのだ。つまりタンパク質はDNAの情報に基づいてアミノ酸から組み立てられるのである。
さて、DNAにはどのようにして情報が記録されてるのだろうか?。DNAはヌクレオチドと総称される4種類の簡単な有機物からできているが、この4種類のヌクレオチドの配置がタンパク質の構造を示している。
しかし、以上のようなDNAの配列をそのまま使って、直接、タンパク質が合成されるわけではない。DNAの配列は鋳型となるメッセンジャーRNAにコピーされ、そしてこのDNAの鋳型のヌクレオチドの配列からタンパク質が作られていく。
つまりタンパク質合成の過程は、
DNA→メッセンジャーRNA→タンパク質
という流れで表すことができる。この流れはすべての生物(一部のウイルスを除く)で普遍的なものだ。しかし、例外がある。
DNAの長い長い配列、人間の場合ならヌクレオチド30億個分の配列のなかには、さまざまな情報や機能が記録されているが、中には”奇妙な性質をもった配列”がまぎれ込んでいる。そのひとつが数百個のヌクレオチドからできているレトロポゾンだ。
レトロポゾンの配列は面白い機能を持っている。DNA上の”本体”からコピーされた鋳型(つまりメッセンジャーRNA)は、ありうべからざることに再びコピーされてしまう。そのためにメッセンジャーRNAの鋳型、つまり最初のレトロポゾンのまったくの複製が出来上がってしまう。そして、それがふたたびDNAのどこかに入り込んでくるのだ。
つまり、レトロポゾンは数十億個のヌクレオチドが連なる長い長い列の中で勝手に増殖し、どこかへ適当に入り込んで”子孫”を作っていく配列なのである。
ではレトロポゾンのこの奇妙な性質がどうして生物の進化を解析する手がかりになるのだろうか。
例えば、3種類の生物、a種・b種・c種のDNAを比較したとき、そのうちa種とb種がDNAの同じ場所に同じレトロポゾン(仮にレトロポゾンXとする)を持っていたら何が言えるだろうか。
まず言えることは、a種とb種は共に
”DNAの同じ場所に同じレトロポゾン、すなわちレトロポゾンXが入り込んだ”
という経験を過去にしていたことになる。これはどういうことを示しているのだろう。次ぎの2つの可能性が考えられるだろう。
1:”a種の祖先”と”b種の祖先”で別々にレトロポゾンXが入り込んだ。→つまり何十億個ものヌクレオチドが並んでいるDNAの同じ場所に同じものが偶然、別々の種で別々に入り込んだ。
2:a種の祖先とb種の祖先はかつて同じ生物、祖先Xであった。→祖先XのDNAのある場所にレトロポゾンXが入り込んだ。祖先Xはその後、幾つかの子孫を生み出し、a種とb種が生まれた。a種とb種は祖先Xから”同じ場所に同じレトロポゾンXが入り込んでいる”という共通の特徴を受け継いでいる。
さて以上のいずれが正しいのだろう?。そもそもDNAの長い配列のまったく同じ場所に”たまたま”同じレトロポゾンが偶然入り込むなんてことがあるだろうか?。その可能性は0ではないものの、とてつもなく低い、要するに事実上ありえないと考えるのが妥当だろう。
するとa種とb種は共通の祖先Xを持つが、c種は持たない、つまり祖先Xから進化したわけではないことが推測できる。その関係は以下のように描くことができるだろう。
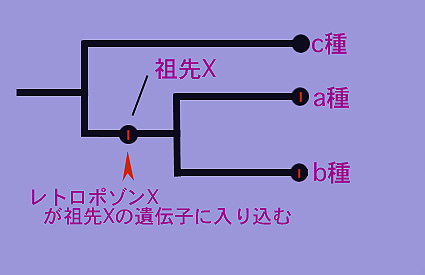
”ある特定のレトロポゾンを共有する”複数の種は祖先を共有することが示唆される。以上の本文で述べたように、
a種とb種が共通のレトロポゾンXを持ち、その一方でc種がレトロポゾンXを持たない、
のならばこの図のような系統関係が描き出せる。
このような手法でクジラと偶蹄類の類縁関係を解析するとどうなるだろうか。97年に発表された島村たちの報告は数種類のクジラと代表的な偶蹄類のレトロポゾンを比較したものだが、そこで明らかにされたのはクジラとウシ・カバは、ラクダやブタが持っていない”固有のレトロポゾン”を幾つも共有するという事実だった。つまり、ウシやカバはラクダやブタよりもクジラに近い動物であり、クジラは偶蹄類の仲間なのだ。
岡田研究チームによるこの成果は古生物学者のこれまでの見解とはまっこうから対立するものだ。一部の研究者は島村たちの結果を
”DNAの類似だけに基づいた系統解析がまた出た”
という程度にとっていたふしがあるが、レトロポゾンを用いた系統解析は、分子系統学のたいてい手法とは違うし、従来なかった新しいアプローチである。
注:分子系統学では幾つもの解析手段があり、そのひとつはレトロポゾンや分岐学と同じ最節約法だが、多くの解析手段は類似度に基づいている。一般向きには上島 96 にまとめられている。
むしろ彼らの手法は、形態から進化を考える分岐学と同じ原理に基づいている。どちらも生物が共通して持つ”特徴(=形質と呼ばれる)”が系統関係を明らかにする証拠と考えているからである。違うのは分岐学で注目する特徴は骨や筋肉、神経の配置などといった”形態”であるのに対して、岡田研究チームが注目するのはレトロポゾンであるということだけでしかない。
しかし、レトロポゾンは形態に対して非常に有利な点がある。
まず、a種とb種に見られる特徴が同じなのか否なのか、それが明瞭であるという点だ。なぜなら比較したレトロポゾンが同じものかどうかは、レトロポゾンのヌクレオチドの配列を見ればよい。また同じ場所にあるかどうかも周囲のDNAのヌクレオチドを見れば一目瞭然である。
また、レトロポゾンには形態でしばしば見られる収斂や”特徴が二次的に失われる”ということがない。同じレトロポゾンがDNAの同じ場所に独立して入り込んだりすることは、限り無く0に近いし、入り込んだレトロポゾンだけが切り出されてなくなるということもありえそうにない。
ようするにレトロポゾンは理論上、収斂や逆転、主観的な同定といったことがほとんどありえない形質と考えてよいというわけだ。
ではこの手法にはなんの問題もないのだろうか。岡田教授によると、幾つかの問題点、例えば祖先多形とよばれる現象に影響を受けることなどがありうるが、島村たちが行なった解析にはそうした際に起こる現象、レトロポゾンの分布の矛盾やレトロポゾンが検出されないという結果はでなかったという。
また、岡田研究グループによるレトロポゾンを用いた系統解析にはこれまでにサケのものがある。そこでは今まで行なわれてきた形質および分子系統学のものと同じ結果が得られている。この事実は理論的な予想からだけでなく、実際の結果から考えてもレトロポゾンによる解析の妥当性を疑う理由がないことを示している。(岡田 98 pp70~92)
ところがサケの場合と違って、クジラについては形態に基づいた結果と矛盾する結論が出ている。このように異なる手法で異なる結論が出ることは珍しいことではないし、また無意味なことでもない。矛盾から形態の評価を考え直したり、遺伝子の解析で考えた前提条件を見直す、よい機会となる。ではクジラにおけるこの矛盾はどう考えるべきなのだろう。
幾人かの研究者は独立してレトロポゾンがDNA上の同じ位置に入り込む可能性を指摘するが、なぜそのような非常に可能性が低いことが起こるのか、その機構について具体的な提案をした人はどうも見当たらない。
要するに、
”形態に基づく分岐図が正しいと仮定した場合、レトロポゾンが独立に同じ場所に入り込んだと考えるべきである”
と言った人はいるが、それ以上のことを語った研究者はいまだにいないようだ。このことは”正しいと仮定することはできるが、その根拠は存在しない”ということのように思える。
それでは逆に考えてみよう。少なくともレトロポゾンが独立して同じ場所に入り込む可能性、つまり数十億のヌクレオチドのまったく同じ領域に入り込む可能性は事実上0であると考えてよい。
つまり岡田研究チームの系統樹が正しいと仮定する根拠は以上のように存在する。すると、岡田研究チームの描いた系統図と、形態の観察から作られた系統図の不一致はむしろ形態の評価に問題があることを示しているのかもしれない。
ではクジラと偶蹄類を改めて観察すれば共通する特徴を見つけられるのだろうか。しかしながらこれは非常に難しい作業だ。なぜなら、クジラは水中生活に適応するために身体があまりにも変型してしまったので、比べるのがとても難しい。
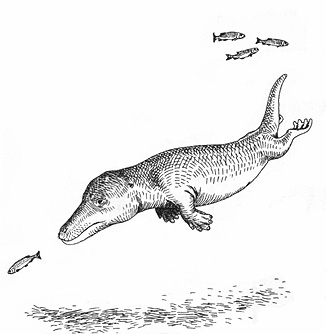
初期のクジラ、アンブロケタス。アンブロケタスは非常に原始的なクジラだが、以下に本文で述べるパキケタスの方がより原始的である(パキケタスはアンブロケタスも含むすべてのCetacea の外群となる)。
ともあれ、保存のよいアンブロケタスの化石は初期のクジラの進化について多くのことを教えてくれる。まず彼らは外見上、現在のクジラとずいぶん異なる姿をしていたことが分かる。また、アンブロケタスは半水棲の生活、つまりワニのような生活をする動物だったらしい。
例えば、後足などそのさいたるもので、現生のクジラでは痕跡がわずかに残っているだけだ。また、クジラは水中で音を聞くために内耳も含めて耳の構造が大きく変わってしまっている。ところがやっかいなことに偶蹄類の重要な特徴とされるものは後足のかかとの骨(距骨:きょこつ)や足の甲の骨(中足骨:中足骨)であったり内耳の骨であったりする。だから少なくとも骨をみる限りでは、現生のクジラと偶蹄類を比較しようがないのである。
しかし、化石を調べると原始的なクジラの中には特殊化がさほど進んでいないために、ある程度比較が可能なものがある。
90年、ミシガン大学のギングリッチたちはまだ比較的大きな後足を残していた初期のクジラ、バシロサウルスの後足に偶蹄類的な特徴があることを報告している。(Gingerich et al. 90)
ギングリッチたちによると、バシロサウルスの後足はパラクソニック・フート(=Paraxonic foot)で第3中足骨と第4中足骨が最も大きく、長くなっていて、2本の骨は脚の中心を平行に伸びている。これは偶蹄類のものと似た構造だ。とはいえ、彼らが指摘するようにこの構造は偶蹄類だけでなく、メソニックス類にも見られるものだ。
一方、オハイオ大学のテゥビッセン教授は94年の論文の中でクジラと偶蹄類、およびメソニックス類とでは運動の仕方が全く異なるのに、脚に同じパラクソニックな構造が見られることに注目している。(Thewissen. 94)
またテゥビッセン教授は内耳にある3つの骨の1つ、キヌタ骨の特徴に注目する。なぜなら、偶蹄類のキヌタ骨は極めて独特のものなのだが、最も原始的なクジラ、パキケタスのキヌタ骨は偶蹄類のそれと似ているからだ。(Thewissen & Hussain 93. )これはクジラと偶蹄類の類縁関係、それもクジラが偶蹄類に含まれる動物であることを暗示している。もっとも、キヌタ骨の資料は現生の偶蹄類とクジラ、そしてパキケタスしかないために、この比較が決定的というわけでもない。(Thewissen 94)
しかしながら、以上のような化石証拠や軟組織に見られる共通点から、テゥビッセン教授は形態と分子の系統樹が一致しないのは形態のデーターが大量に失われていることが原因ではないか、と述べている。(Thewissen. 94)
さらに98年、テゥビッセン教授はパキスタンから原始的なクジラのものとおもわしき距骨を見つけた。先に検討された中足骨やキヌタ骨と同様、この発見は偶蹄類とクジラの類縁関係を暗示する一方で、いささか人を当惑させずにはいられない。なぜなら、この原始的なクジラの距骨は偶蹄類とまったく同じではなかったからだ。とはいえ、興味深いことにこのクジラの骨が偶蹄類に固有の特徴を持っているのもまた事実であり、決定打ではないにしても、またもや偶蹄類とクジラの関係を暗示させる証拠が見つかったわけである。(Thewissen et al 98)
さてここでレトロポゾンによる系統解析に話を戻そう。島村たちの研究に続いて、岡田研究チームの二階堂雅人たちはレトロポゾンの探索を続け、99年、偶蹄類の中でもカバがクジラに最も近縁であることを示した。(Nikaido et al. 99)
二階堂たちの報告によると、カバとクジラの類縁性は4つのレトロポゾンを共有することで支持される。また、近年、行なわれた分子系統学による研究結果もクジラが偶蹄類に含まれ、またクジラがカバと最も近い関係にあることを示している。(Murphy et al 01)
同じ遺伝子に基づいた系統解析とは言っても、レトロポゾンを用いた二階堂たちの報告と、他の分子系統学者の研究とはまったく異なるアプローチから得られたものだ。
この”一致”は大きい。
なぜならレトロポゾンを用いた解析手段の妥当性は理論的に予想されているが、さらに加えて異なる解析手段が同じ結論を示したのである。異なるアプローチが同じ結論を導き出すなら、その結論の妥当性は理論にとどまらず、いやがおうにも高くなる。
さて、以上の事柄を踏まえると、私達はクジラや偶蹄類の形態をもう一度再評価するべききではないかと感じざるおえない。実は軟組織の研究、つまり解剖学の視点からすると偶蹄類とクジラの類縁を支持する証拠が幾つもある。(例えばThewissen 94 にまとめられている)これはなにを示しているのだろうか。
最近、国立科学博物館の甲能直樹研究官は、偶蹄類とクジラ類の軟組織に見られる特徴の幾つかは、二階堂たちが明らかにした”系統樹”に当てはめることができることを示した。(甲能 00)
もちろん、これは系統解析に基づいていないから、二階堂たちの系統を支持する共有派生形質ではない。また、甲能研究官が断っているようにまったく整合しない形質が、一方であることも事実である。
とはいえ、
”二階堂たちの系統を支持する共有派生形質のように並ぶ形質”
が複数存在するということは興味深い。
以上のことを踏まえると、私達は今一度、クジラやカバ、偶蹄類の形態を見なおす必要があるのだろう。しかし、その一方ではクジラやカバ、および偶蹄類の基礎的な解剖学の知見すら不十分であるという重い現実が立ちはだかっている。
つまるところ前世紀に終わったと思い込んでいた”解剖”という基本的な作業を見直すことさえ、今、必要とされているのだろう。そしてこのことは”げんなりする作業”であるというより、古くて新しい学問に未来があるということのように思われる。
レトロポゾンが突き付けた衝撃は、今や忘れられていた学問の再検討にまで余震がおよぼうとしている。しかし以上に述べたような理由から、私達はそのことをむしろ喜ぶべきかもしれない。
見直しから私達はいかなる知見に達することができるだろうか?